| |
Geographical Speciation
A barrier restricts
gene flow between populations so that they evolve separately and
eventually become different species. The barrier may break down
and the isolates may again interact but they do not interbreed.
(Rosenzweig,
1995).
The dynamics of speciation
will depend on two processes: 1) the rate at which geographical
isolates are formed and 2) the rate at which these isolates evolve
into separate species.
1. Rates of isolate
formation are influenced by spatial factors
a) geographical
circumstance: archipelagos and mountain ranges are very
effective isolating barriers. Once something does overcome the
barrier to colonize these areas, speciation tends to occur relatively
rapidly. Therefore, mountains and archipelagos will tend to
increase rates of speciation there.
b) geographical
range: size of the geographical range of an organism makes
it more or less likely to include a barrier. A range can be
too small, making it unlikely that a barrier will pass through
it. A very large range may wholly encompass a barrier, so that
individuals (or propagules such as pollen or spores) can pass
around it. Therefore, intermediate-sized ranges are most likely
to be divided by a barrier (knife-shaped).
2. Rates of speciation
following isolate formation are influenced by two factors:
a) sexual divergence:
the isolates may evolve different mating behaviour, for example,
by flowering at different times of the year
b) ecological
divergence: natural selection will cause different isolates
to evolve differently because no two places are exactly alike.
Speciation is more
likely to occur in large populations than in small populations
because they contain more variation. This variation cannot be
selected effectively, however, if the population is completely
interbreeding. The most likely situation for divergence to occur,
therefore, is when a a relatively small and unrepresentative group
(a "propagule") is split off, or isolated, from a large ("parent")
population.
THE
evolution of Darwin's finches on the Galapagos Islands is a dramatic
exam ple
of geographic speciation. It is believed that the 13 species of
Darwin's finches that are found on the Islands descended from
an ancestral pair of South American finches that landed there
accidentally over 100 000 years ago. This pair found an area free
of predators and probably adapted to the various unfilled niches.
For instance, one finch population evolved a longer bill and the
facility for using sticks to prod insect holes in cactus, and
over time evolved into the Woodpecker finch. Other finches evolved
thicker bills for eating the large seeds of the prickly pear cactus,
and became the Large ground finch; smaller thick bills were ideal
for eating small seeds; while other bills and habits adapted themselves
to insect predation. Gradually, a number of such variations led
to the radiation of 13 different species from 1. However, this
classification of "species" is tenuous, as some of the species
are thought to be able to interbreed. ple
of geographic speciation. It is believed that the 13 species of
Darwin's finches that are found on the Islands descended from
an ancestral pair of South American finches that landed there
accidentally over 100 000 years ago. This pair found an area free
of predators and probably adapted to the various unfilled niches.
For instance, one finch population evolved a longer bill and the
facility for using sticks to prod insect holes in cactus, and
over time evolved into the Woodpecker finch. Other finches evolved
thicker bills for eating the large seeds of the prickly pear cactus,
and became the Large ground finch; smaller thick bills were ideal
for eating small seeds; while other bills and habits adapted themselves
to insect predation. Gradually, a number of such variations led
to the radiation of 13 different species from 1. However, this
classification of "species" is tenuous, as some of the species
are thought to be able to interbreed.
Competitive
speciation
Competitive speciation
occurs when one portion of a population exploits a new ecological
niche or opportunity (food, life history attribute, habitat, etc.)
that was previously unexploited and becomes sufficiently different
as to be considered a new species. This is the most controversial
mode of speciation
(Rosenzweig, 1995).
The place where a population
lives may contain two (or more) different kinds of resource, for
example, two species of food plant. Some individuals may use one
plant more effectively, and some the other plant. These specialists
are likely to be more successful than individuals who are not
as effective in using either plant. Specialists who mate among
themselves will be exceptionally successful, because their offspring
are likely to inherit their specialization. The evolution of appropriate
mating preferences may then lead to the appearance
of two separate groups, who in time become so strongly isolated
that they become different species.
The problem with this
explanation is that random mating among the different phenotypes
and genetic recombination break up any adaptive combinations of
genes faster than they can be selected. For sympatric speciation
to work, therefore, some strong force has to hinder recombination.
In other words, something would have to prevent an individual
suited to environment A from mating with an individual suited
to some environment in between A and B, even though they are the
same species and in the same location. (Rosenzweig,
1995)
STRONG
evidence for competitive effects was shown in a 1969 paper by
Guy Bush who studied the fruit flies of the genus Rhagoletis.
Prior to European arrival in North America ,
R. pomonella fed exclusively on hawthorn and R. indifferens
on native pin cherry. Each of these species has now formed a new
host race, the former adapted to domesticated apple and the latter
fed on cherries. Both these fruits were introduced by Europeans
and both flower at slightly different times from their native
counterparts; therefore the races that became adapted to the domesticated
fruits had a different reproductive cycle from those that fed
on native fruit. This temporal difference has resulted in races
of the same species in the same location that are reproductively
isolated - one of the main criteria defining different species.
(Bush,
1969) ,
R. pomonella fed exclusively on hawthorn and R. indifferens
on native pin cherry. Each of these species has now formed a new
host race, the former adapted to domesticated apple and the latter
fed on cherries. Both these fruits were introduced by Europeans
and both flower at slightly different times from their native
counterparts; therefore the races that became adapted to the domesticated
fruits had a different reproductive cycle from those that fed
on native fruit. This temporal difference has resulted in races
of the same species in the same location that are reproductively
isolated - one of the main criteria defining different species.
(Bush,
1969)
Polyploidy
 Most
familiar organisms have two sets of chromosomes, inheriting one
set from each parent. Such organisms are called diploid. Polyploid
individuals have more than two sets. They arise through cytological
irregularities during cell division or through the fusion of abnormal
gametes. Once formed, they are often sexually isolated from their
parent population. For example, a tetraploid individual (having
four sets of chromosomes) forms diploid gametes. When these fuse
with the haploid gametes produced by normal individuals, they
give rise to triploid progeny (3 sets of chromosomes), which are
sterile. This is why polyploidy can result in instant, or abrupt,
speciation. Most
familiar organisms have two sets of chromosomes, inheriting one
set from each parent. Such organisms are called diploid. Polyploid
individuals have more than two sets. They arise through cytological
irregularities during cell division or through the fusion of abnormal
gametes. Once formed, they are often sexually isolated from their
parent population. For example, a tetraploid individual (having
four sets of chromosomes) forms diploid gametes. When these fuse
with the haploid gametes produced by normal individuals, they
give rise to triploid progeny (3 sets of chromosomes), which are
sterile. This is why polyploidy can result in instant, or abrupt,
speciation.
Which type of speciation
is most prevalent?
Polyploid series of
related species are easy to identify and can be common, especially
among plants. More generally, most speciation probably requires
isolation, followed by divergence: good examples of sympatric
speciation are hard to find. (Rosenzweig,
1995)
Neutral
processes that regulate species diversity
Neutral processes are
those that occur independently of any differences among species,
as though the species were genetically identical.They will, therefore,
affect diversity regardless of the ecological characteristics of
a region. For example, there is a continual rain of seeds and spores
onto the soil, and which species happen to land in a site suitable
for growth is largely a matter of chance.
Immigration
Immigration
provides a continual source of new diversity for a region. How important
it is depends on the balance between the number of propagules that
come from outside and the number produced by resident individuals.
If the area is large (a few square kilometers), most young individuals
will be recruited from the resident population, but in small areas
(a few square meters), reproduction by residents may be overwhelmed
by immigration. Thus the importance of immigration increases
as the size of the area decreases.
Some
organisms are dispersed much more broadly than others.
The very small spores of ferns, for example, may be carried by
wind for hundreds of kilometers from their parental site. The
seeds of plants such as dandelions and poplars are much larger
but have special devices to facilitate wind transport. Marine
creatures such as corals and starfish have larvae which are carried
for great distances on ocean currents. Immigration will be much
more important in such creatures than it will be in oak trees
or land snails, for example, which produce larger propagules with
no special devices to ensure long-distance dispersal.
The effectiveness of
immigration in providing new recruits to an area is seen dramatically
after a natural disaster destroys all life in an area. After the
devastation of a volcanic eruption, for example, plants and animals
quickly return: first the groups with effective long-distance
dispersal, and later, those who disperse more slowly but are better
competitors once they arrive. (Rosenzweig,
1995)
Extinction
Extinction
of a species or a population will occur for one of two reasons:
as a result of accidents (environmental fluctuations) or because
of population interactions.
a) accidents:
events that trigger extinctions for no predictable reason -
volcanos, rising sea level, an ice storm, any environmental
circumstance that wipes out an ecological niche.
b) population
interactions that are not neutral processes: predation and
competition can result in negative growth rate and ultimately,
extinction. However, on their own, predation and competition
rarely cause extinctions directly; they cause population densities
to become very low and then a random accident may drive the
vulnerable population to extinction.
The probability that
enviornmental or population fluctuations will cause an extinction
depend on how abundant the organism is and how large its range
is.
a) abundance:
if the chance that any given individual will die in a given
period of time is p, then the chance that all individuals in
a population of size N will die within that same period of time
is pN. If the population is large, the probability that this
will happen is very small: for example, if p = 0.5 in a given
year, then the probability that all individuals in a population
of N = 1000 will die at the same time is so small that it is
unlikely to occur in a billion years. If, on the other hand,
N = 10, the population is likely to become extinct within a
thousand years, a relatively short period, and certainly much
shorter than the time necessary to produce a new species. Small
populations are thus at high risk of chance extinction.
b) range:
disturbances that kill all the individuals in a given area happen
all the time. Smaller and more localized disturbances are more
frequent than large and widespread disturbances - treefalls
are more frequent than forest fires, landslips more common than
earthquakes. A species that is restricted to a few small sites
is therefore at higher risk of being extinguished by an environmental
fluctuation than one that occurs at many sites over a large
area. (Rosenzweig,
1995)
Theory of Island
Biogeography
The equilibrium
theory of island biogeography states that the number of species
on a given island is regulated by the balance between immigration
of new species and extinction of species.
The
total number of species found on an island depends on the size
of the island and the distance from the source of immigrants or
propagules (mainland or other islands). The smaller the island,
the higher the probability that the population can be wiped out
by random fluctuations in size, environmental conditions and mortality.
An island that is near to the source population will receive more
immigrants than one that is far away because more individuals
will be able to cross the barrier (water, or other hostile habitat).
Therefore, we expect to find more species on larger islands that
are closer to the mainland or to other islands and fewer species
on small distant islands.
By extension, the same
principle applies to any stretch of land, whether it is separated
by land or not. And, of course, one can readily view lakes as
islands of water surrounded by land. The theory, therefore, is
broadly applicable to all sorts of situations.
Island biogeography
theory is a neutral theory, because species are assumed to have
the same rates of extinction and immigration. It leads to a characteristic
species diversity on islands of given size and isolation as the
result of a dynamic equilibrium between the processes of extinction
and re-colonization. We can therefore use the theory to predict
patterns of species diversity.
(MacArthur
and Wilson, 1963; Huston,
1996, Ch.4; Rosenzweig,
1995, p.220-263)
|


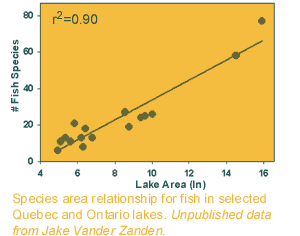 the
bigger the area sampled, the more species found.
the
bigger the area sampled, the more species found. 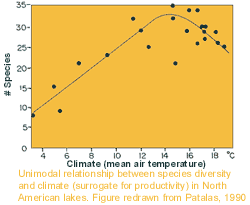 But the pattern is usually more complicated than this. In many
systems, the relationship between primary productivity and diversity
has been shown to be unimodal, or hump-shaped: diversity is highest
at intermediate levels of productivity. Such a pattern has occurred
at regional scales in many biomes and for many groups of animals
and plants, including desert rodents, tropical mammals, marine
benthic communities and megafauna, freshwater plankton, montane
ferns and bryophytes.
But the pattern is usually more complicated than this. In many
systems, the relationship between primary productivity and diversity
has been shown to be unimodal, or hump-shaped: diversity is highest
at intermediate levels of productivity. Such a pattern has occurred
at regional scales in many biomes and for many groups of animals
and plants, including desert rodents, tropical mammals, marine
benthic communities and megafauna, freshwater plankton, montane
ferns and bryophytes.  Disturbance
regimes, therefore, play an important role in the maintenance
of regional species diversity, mainly by preventing, or slowing
down competitive exclusion by one dominant species by
Disturbance
regimes, therefore, play an important role in the maintenance
of regional species diversity, mainly by preventing, or slowing
down competitive exclusion by one dominant species by 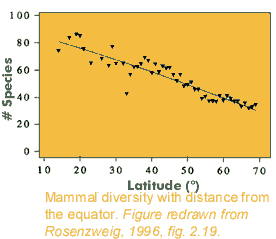 Latitudinal Gradients:
Scientists have observed that species diversity declines with
increasing distance from the Equator, either north or south.
There is no doubt that the tropics are very rich in species
diversity. And this pattern is ancient; it has existed for thousands,
if not millions, of years, across many taxa, from trees to fossil
foraminifera.
Latitudinal Gradients:
Scientists have observed that species diversity declines with
increasing distance from the Equator, either north or south.
There is no doubt that the tropics are very rich in species
diversity. And this pattern is ancient; it has existed for thousands,
if not millions, of years, across many taxa, from trees to fossil
foraminifera. 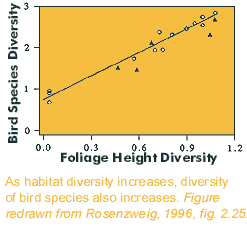 Habitat
Variety: The more variable the habitat, the greater the
species diversity within it. This pattern was offered as one
of the reasons why there are more species in a bigger area (more
area covers a greater variety of habitat).
Habitat
Variety: The more variable the habitat, the greater the
species diversity within it. This pattern was offered as one
of the reasons why there are more species in a bigger area (more
area covers a greater variety of habitat).